
Текущее название
| Index Fungorum | Heterobasidion annosum (Fr.) Bref. | |
| MycoBank | Heterobasidion annosum (Fries) Brefeld |
Систематическое положение
Fungi, Basidiomycota, Agaricomycetes, Russulales, Bondarzewiaceae, Heterobasidion
Этимология видового эпитета
Annosus, a, um, многолетний, старый.
Синонимы
- Polyporus annosus Fr., Syst. mycol. (Lundae) 1: 373 (1821)
- Boletus annosus (Fr.) Spreng., Syst. veg., Edn 16 4 (1): 278 (1827)
- Trametes annosa (Fr.) G.H. Otth, Mitt. naturf. Ges. Bern 531-552: 73 (1864)
- Fomitopsis annosa (Fr.) P. Karst., Revue mycol., Toulouse 3 (no. 9): 18 (1881)
- Fomes annosus (Fr.) Cooke, Grevillea 14 (no. 69): 20 (1885)
- Placodes annosus (Fr.) Quél., Enchiridion Fungorum in Europa media et praesertim in Gallia Vigentium: 172 (1886)
- Pycnoporus annosus (Fr.) P. Karst., Finl. Basidsvamp. (no. 11): 12 (1898)
- Scindalma annosum (Fr.) Kuntze, Revis. gen. pl. (Leipzig) 3 (3): 518 (1898)
- Ungulina annosa (Fr.) Pat., Essai Tax. Hyménomyc. (Lons-le-Saunier): 103 (1900)
- Friesia annosa (Fr.) Lázaro Ibiza, Revta R. Acad. Cienc. exact. fis. nat. Madr. 14 (9): 588 (1916)
- Boletus cryptarum Bull., Herb. Fr. (Paris) 10: tab. 478 (1790)
- Poria scutata Hoffm., Veg. Herc. subterr. 12: 13 (1794)
- Polyporus batschii b scutatus Pers., Mycol. eur. (Erlanga) 2: 85 (1825)
- Boletus epiphegus Schumach., Enum. pl. (Kjbenhavn) 2: 389 (1803)
- Trametes scutata (Pers.) Harz, K. svenska Vetensk-Akad. Handl., ser. 3: 135 (1848)
- Polyporus subpileatus Weinm., Syll. Pl. Nov. Ratisb. 2: 102 (1828)
- Polyporus scoticus Klotzsch, Brit. Fung., Phycom. Ustil. (London) 3: 142 (1836)
- Polyporus makraulos Rostk., in Sturm, Deutschl. Fl., 3 Abt. (Pilze Deutschl.) 4: 113 (1838)
- Trametes radiciperda R. Hartig, Wichtige Krankheiten der Waldbäume. Beiträge zur Mykologie und Phytopatologie für Botaniker und Forstmänner (Berlin): 62 (1874)
- Polystictoides fuscus Lázaro Ibiza, Revta R. Acad. Cienc. exact. fis. nat. Madr. 14 (11): 755 (1916)
- Polyporus marginatoides E.H.L. Krause, Arch. Freunde NatGesch. Mecklenb. 3: 54 (1928)
- Cunninghamella meineckella A.J. Olson, Phytopathology 31: 1076 (1941)
Другие названия: Корневая губка.
Таксономия
Мощные фитопатогенные свойства и масштабный урон, который Heterobasidion annosum наносит лесному хозяйству, сделали его объектом пристального и долгосрочного внимания микологов и фитопатологов. Именно поэтому таксономическая история у него весьма обширна и запутанна. Имеет смысл остановиться только на самых значимых этапах:
- Изначально вид описал в 1821 году Элиас Магнус Фриз (Elias Magnus Fries; 1794 – 1878) в своей знаковой работе «Микологическая система», указав основным субстратом берёзу и дав ему имя Polyporus annosus Fr [1].
- В 1874 году немецкий лесовод и миколог Роберт Гартиг (Robert Hartig; 1839 – 1901) вновь описал этот вид, назвав Trametes radiciperda R. Hartig, указав, что грибом поражаются такие породы, как сосна, можжевельник, бук и боярышник [6]. Именно это название использовалось во многих ранних публикациях, особенно в Германии.
- В 1879 году финский миколог-систематик Петер Адольф Карстен (Petter Adolf Karsten; 1834 – 1917) перевёл Polyporus annosus Fr. В род Fomes [7]. Эта комбинация оставалась незамеченной, пока в 1885 году английский ботаник и миколог Мордехай Кук (Mordecai Cubitt Cooke; 1825 – 1914) не повторил перенос [8]. Имя Fomes annosus (Fr.) Cooke было наиболее часто используемым в литературе до конца 1970-х годов.
- В 1881 году Карстен описал новый род – Fomitopsis P. Karst. [9], а чуть позже, в том же году, включил в него и Fomes annosus [10]. Общее описание рода получилось довольно расплывчатым. Например, Маринус Антон Донк (Marinus Anton Donk; 1908 – 1972) считал эту комбинацию недействительной [11]. Первое описание комбинации Fomitopsis annosa (Fr.) Bond. & Singer, соответствующее Кодексу ботанической номенклатуры, опубликовали Аполлинарий Семёнович Бондарцев (1877 – 1968) и Рольф Зингер (Rolf Singer; 1906 – 1994) в 1941 году. Это имя широко использовалось в литературе, издававшейся в Советском Союзе до 1980-х годов [12].
- В 1886 году Люсьен Келе (Lucien Quélet; 1832 – 1899) перевел этот вид в род Placodes [13].
- В 1889 году немецкий миколог Юлиус Оскар Брефельд (Julius Oscar Brefeld; 1839 – 1925) описал новый род – Heterobasidion, а в его рамках – новую комбинацию Heterobasidion annosum (Fr.) Bref [2].
- В 1900 году французский фармацевт и миколог Нарсис Теофиль Патуйяр (Narcisse Théophile Patouillard; 1854 –1926) объединил этот вид с родом Ungulina, типовым видом которого, как и рода Placodes, является Fomes fomentarius, что сделало роды Ungulina и Placodes избыточными синонимами рода Fomes [14].
Помимо перечисленных выше, для комплекса Heterobasidion annosum использовалось множество других названий, которые были достаточно быстро забыты.
Новейшая история немного динамичней. Ближе к окончанию XX века стало очевидно, что H. annosum это комплекс видов. Большие и значимые шаги в попытке разобраться с этим сделал финский миколог и фитопатолог Кари Корхонен (Kari Korhonen). На основе экспериментов по спариванию большого количества культур он разделил H. annosum на две группы интерстерильности. Эти группы получили обозначения «S» и «P» и отличаются среди прочего в предпочтениях к субстрату/хозяину. Группа «S» встречается в основном на ели европейской, но также уничтожает саженцы сосны обыкновенной. Группа «P» наносит вред сосне обыкновенной, но также встречается на некоторых других древесных породах, включая и европейскую ель [40]. В 1990 году была выявлена третья группа интерстерильности, получившая обозначение «F». Эта группа приурочена к пихте белой (Abies alba). Тесно связана с европейской группой «S», но сексуально несовместима с ней [41]. В 1998 году в огромном труде посвящённом целиком корневой губке «Heterobasidion annosum. Biology, Ecology, Impact and Control», Niemelä T. и Korhonen K. присвоили этим группам самостоятельные видовые имена: «S» – Heterobasidion parviporum Niemela & Korhonen; «P» – Heterobasidion annosum (Fr.) Bref.; «F» – Heterobasidion abietinum Niemela & Korhonen [15].
В 2010 году выяснилось, что североамериканские сборы делятся на две взаимостерильные группы, несовместимые с европейскими; они получили названия гетеробазидион западный (Heterobasidion occidentale Otrosina & Garbelotto) и гетеробазидион нерегулярный (Heterobasidion irregulare Garbelotto & Otrosina) [42].
В 2021 году группа азиатских исследователей во главе с профессором Юй-Чэн Даем (Yu-Cheng Dai) провели глобальную молекулярно-филогенетическую ревизию рода Heterobasidion [43], по результатам которой было описано три новых вида из Юго-Восточной Азии: гетеробазидион Арманда (H. armandii) и гетеробазидион подостровной (H. subinsulare) из китайской провинции Юннань, и гетеробазидион подмелкопоровый (H. subparviporum) из провинции Хэбэй, а ранее описанные виды претерпели ряд таксономических транзакций, в результате которых на данный момент авторы выделяют пятнадцать видов в трёх комплексах:
- комплекс H. annosum «F», включающий виды, ранее относимые к группе H. annosum «S»;
- комплекс H. annosum «P», включающий два вида, поражающих в основном сосны — европейский H. annosum s. str. и североамериканский H. irregulare;
- комплекс H. insulare включает десять видов, ассоциированных с различными хвойными породами (отдел Pinophyta): пихтами (Abies), араукариями (Araucaria), лиственницами (Larix), соснами (Pinus), лжелиственницами (Pseudolarix) и псевдотсугами (Pseudotsuga). В этот комплекс входит и гетеробазидион араукариевый (H. araucariae), который встречается в Южном полушарии.
Восемь видов, отмеченных в Гималаях, указывают на возможное происхождение общего предка Heterobasidion из Азии.








Габитус
- Гименофор: Трубчатый, пористый
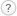
Плодовое тело
Плодовые тела многолетние, разнообразной, часто неправильной формы, в зависимости от возраста и расположения на субстрате могут быть тонкие, корковидные инкрустирующие субстрат, кожисто-деревянистые, раковинообразные, полностью распростертые, распростерто-отогнутые, копытообразные. Размеры, как правило, не превышают 15 см в наибольшем измерении; плодовые тела часто сливаются друг с другом, образуя длинные консоли или черепитчатые полки до 40 см шириной. Поверхность шляпок неровная, концентрически-бороздчатая, складчатая, у молодых экземпляров опушённая, к зрелости покрывается тонкой бурой или каштановой, иногда почти чёрной коркой. Край белый, ровный или волнисто-лопастный, в нижней части стерильный. Ткань у молодых грибов беловатая, по текстуре похожа на мягкую пробку, у старых – более жёсткая и деревянистая, коричневая. Запах характерно душистый, смолистый, интенсивный, иногда настолько, что плодовые тела получается обнаружить по запаху; часто описывается как запах скипидара, но по нашим собственным ощущениям, он очень похож на запах маслят, таких как маслёнок зернистый (Suillus granulatus) или маслёнок обыкновенный (Suillus luteus).
Гименофор трубчатый. Трубочки ежегодно неравномерно нарастают слоями по 2 – 7 мм толщиной, одного цвета с мякотью. Поры круглые или угловатые, иногда неправильной формы, вначале с толстыми, с возрастом утончающимися краями, 2 – 3 (4) шт. на 1 мм.
Микроскопия
Базидиопоры 4.5 – 6.2 × 4 – 5.2 мкм, от почти шаровидной до яйцевидной формы, гиалиновые, мелкошиповатые [5].
Конидии в микроскоп почти неотличимы от базидиоспор, имеют более гладкую поверхность с шипиками 0.045 – 0.02 мкм высотой и менее выраженным, но более длинным центрально-симметричным апикулюсом длиной в среднем 0.56 мкм [30].
Базидии 16 – 22 × 5 – 7 мкм, булавовидные, 4-споровые, без пряжки в основании [5].
Цистиды или другие гимениальные стерильные структуры отсутствуют [5].
Гифальная система димитическая: генеративные гифы гиалиновые, тонкостеные, с редкими ветвлениями, 2.5 – 5 мкм в диаметре, перегородки без пряжек; скелетные гифы гиалиновые, толстостенные, без перегородок, 3 – 5.5 мкм в диаметре; гифы трамы не отличаются от скелетных [5].
Экология и распространение
Растёт на живой и мёртвой древесине хвойных пород, преимущественно на соснах (Pinus), в частности, на cосне обыкновенной (Pinus sylvestris). Редко можно встретить на деревьях лиственных пород (Acer, Alnus, Betula, Carpinus, Corylus, Crataegus, Fagus, Fraxinus, Lonicera, Prunus, Pyrus, Robinia, Sambucus, Salix, Sorbus, Ulmus). Плодовые тела образуются на выступающих из почвы корнях, в комлевой части стволов, на погребённой древесине, на пнях и крупном валеже.
Распространён по всей умеренной зоне северного полушария. Опасный высокоспециализированный возбудитель корневой гнили сосны обыкновенной (Pinus sylvestris L.). В местах широкого распространения вызывает массовую гибель деревьев.
Гриб производит как конидиоспоры, так и базидиоспоры. Первичная инфекция обычно происходит при помощи базидиоспор, которые поражают свежие раны на стволах и корнях деревьев [29][31]. Есть сообщения, что конидиоспоры присутствуют в воздухе [16], но их роль в распространении гриба в природе пока не ясна. Возможно они важны для передачи на короткие расстояния или взаимодействуют с насекомыми, питающимися корнями [17].
В регионах с умеренным климатом базидиоспоры выделяются в основном летом, когда пни наиболее восприимчивы к инфекциям. При температурах ниже 5° C гриб приживается гораздо реже, либо из-за отсутствия базидиоспор, либо из-за неспособности расти при низкой температуре, либо из-за сочетания этих факторов [18]. Также заражения не происходит в периоды, когда температура субстрата превышает 35° C [19]. После прорастания базидиоспор мицелий может жить на пнях и валеже в течение многих лет, не вызывая заболеваний живых деревьев, но может быть и значительно более агрессивным [20]. После колонизации пней гриб распространяется через корневые контакты инфицированных деревьев к здоровым.
H. annosum некротрофно растёт в заболони живых деревьев, и со временем быстро разрастается в сердцевине у большинства видов, за исключением сосен, экстрактивные вещества сердцевины которых фунгистатичны [21]. Скорость распространения сильно варьируется и во многом зависит от типа и возраста древостоя, состава леса и свойств почвы, включая pH. В северных лесах умеренного пояса зарегистрированы средние темпы роста 20 – 50 см в год [22][23].
Было отмечено, что H. annosum способен сохраняться в корневой системе заражённых деревьев в течении десятилетий, легко поражать берёзу, пересаженную на заражённые участки вместо сосны, и эффективно передаваться от одного поколения леса к другому. При этом молекулярные исследования показали, что очаги диаметром до 50 метров представляли собой клон одного и того же экземпляра H. annosum возрастом 35 – 40 лет [24][25][26]. Обезопасить следующее поколение леса на заражённом ранее участке можно дополнительными действиями после санитарной рубки – удалением пней, а также фрагментов корней путём просеивания почвы.[27].
Плодоношение
Плодовые тела можно обнаружить в течении всего года.
Пищевые свойства
Сходные виды
В первую очередь вид сходен с другими представителями рода Heterobasidion. Ниже отметим два европейских вида, хотя не исключено, что масштабное молекулярно-генетическое изучение сибирских и дальневосточных образцов расширит список видов, присутствующих на территории России.
- Гетеробазидион мелкопоровый (Heterobasidion parviporum) в основном поражает ель обыкновенную (Picea abies). Для него характерны мелкие поры (4 – 6 шт. на 1 мм) и хорошо развитое войлочное опушение поверхности шляпок у молодых грибов.
- Гетеробазидион пихтовый (Heterobasidion abietinum) чаще всего встречается на пихте (Abies). Плодовые тела чаще более массивные, толстые, с более округлым, тупым краем, чем у других видов комплекса. На разрезе годовые слои хорошо отграничены друг от друга.
Десять известных видов, встречающихся в Азии, являются сапротрофами, в отличие от североамериканских H. occidentale и H.irregulare и европейских H. abietinum, H. annosum s. str., и H. parviporum; эти пять видов следует считать карантинными, хотя H. irregulare уже отмечался в Италии [43].
- Трутовик плоский (Ganoderma applanatum) в молодом возрасте может напоминать H. annosum. Отличается мякотью коричневого цвета, темнеющим при повреждении гименофором, более правильной формой плодовых тел и своеобразным кисловато-пряным запахом, непохожим на упомянутый выше «запах маслят» или «скипидарный» запах.
Заметки
Наиболее эффективным и экологически безопасным способом борьбы с корневой губкой на данный момент считается использование грибов-антагонистов, в первую очередь флебиопсиса гигантского (Phlebiopsis gigantea), более известного в лесоводческой литературе как пениофора гигантская (Peniophora gigantea). После санитарных рубок вместо раскорчёвки, приводящей к сильному нарушению почвенного покрова, оставшиеся пни инокулируются препаратами живой культуры Ph. gigantea. Этот дереворазрушающий гриб, образующий распростертые плодовые тела серого цвета, растёт значительно быстрее корневой губки и не поражает живые деревья [32][33][34][35].
Ряд исследований указывает на эффективность применения триходермы зелёной (Trichoderma viride) в качестве агента биологического контроля корневой губки, что подтверждается и практическим применением живых культур этого гриба [36][37][38]. Другим исследователям, однако, не удалось подтвердить результаты, полученные в лабораторных условиях, в последующих полевых экспериментах [39].
Материалы по теме
- Fries E. M. Systema Mycologicum. Vol. I. Lund: Ex Officina Berlingiana, 1821. — P. 373. [As Polyporus annosus]
- Brefeld O. Basidiomyceten III. Autobasidiomyceten und die Begründung des natürlichen Systemes der Pilze. // Untersuchungen aus dem Gesammtgebiete der Mykologie. — 1889. — V. 8. — P. 1–305. — P. 154.
- Бондарцева М. А. Порядок Афиллофоровые. Вып. 2. / Определитель грибов России. — СПб.: «Наука», 1998. — 391 с. — С. 225.
- Breitenbach J, Kränzlin F. Fungi of Switzerland. A contribution to the knowledge of the fungal flora of Switzerland. Vol 2. Heterobasidiomycetes, Aphyllophorales, Gasteromycetes. — Lucerne: Verlag Mykologia, 1986. — 412 p. — P. 314.
- Ryvarden L., Melo I. Poroid fungi of Europe. / Synopsis Fungorum. Vol. 31 — Oslo: Fungiflora, 2014. — 455 p. — P. 213.
- Hartig R. Wichtige Krankheiten der Waldbäume. Beiträge zur Mykologie und Phytopathologie für Botaniker und Forstmänner. — Berlin: Verlag von Julius Springer, 1874. — 127 p. — P. 62. [As Trametes radiciperda]
- Karsten P. Symbolae ad Mycologiam Fennicam. // Meddelanden af Societas pro Fauna et Flora Fennica. — 1879. — V. 5. — P. 38. [As Fomes annosus (Fr.) P. Karst.]
- Cooke M. C. Praecursores ad Monographia Polypororum. // Grevillea. — 1885. — V. 14. — P. 20. [As Fomes annosus (Fr.) Cooke]
- Karsten P. Symbolae ad Mycologiam Fennicam. // Meddelanden af Societas pro Fauna et Flora Fennica. — 1881. — V. 6. — P. 9.
- Karsten P. Enumeratio Boletinearum et Polyporearum Fennicarum, Systemate novo Dispositarum. // Revue Mycologique (Toulouse). — 1881. — V. 3 (9). — P. 18. [As Fomitopsis annosa (Fr.) P. Karst.]
- Donk M. A. The generic names proposed for Polyporaceae. // Persoonia. — 1960. — V. 1 (2). — P. 173–302. — P. 217.
- Bondartsev A., Singer R. Zur Systematik der Polyporaceen. // Annales Mycologici. — 1941. — V. 39 (1). — P. 43–65. — P. 55. [As Fomitopsis annosa (Fr.) Bond. & Singer]
- Quélet L. Enchiridion Fungorum in Europa Media et praesertim in Gallia Vigentium. — Lutetiae: Octave Dion, 1886. — 352 p. — P. 172. [As Placodes annosus (Fr.) Quél.]
- Patouillard N. Essai taxonomique sur les familles et les genres des Hymenomycetes. — Lons-le-Saunier, 1900. — 184 p. — P. 103. [As Ungulina annosa (Fr.) Pat.]
- Niemelä T., Korhonen K. Taxonomy of the genus Heterobasidion. Pages 27–33 in: Woodward S., Stenlid J., Karjalainen R., Hüttermann A. (eds) Heterobasidion annosum. Biology, Ecology, Impact and Control. — CAB International, 1998. — 589 p.
- Hsiang T., Edmonds R. L., Driver C. H. Conidia of Heterobasidion annosum from Tsuga heterophylla forests in western Washington. // Canadian Journal of Botany. — 1989. — V. 67 (4). — P. 1262–1266. DOI: 10.1139/b89-164.
- Kadlec Z., Starý P., Zumr, V. Field evidence for the large pine weevil, Hylobius abietis as a vector of Heterobasidion annosum. // Forest Pathology. — 1992. — V. 22 (5). — P. 316–318. DOI: 10.1111/j.1439-0329.1992.tb00799.x.
- Meredith D. S. The Infection of Pine stumps by Fomes annosus and other fungi. // Annals of Botany. — 1959. — V. 23 (3). — P. 455–476. DOI: 10.1093/oxfordjournals.aob.a083672.
- Ross E. W. Fomes annosus in the southeastern United States: relation of environmental and biotic factors to stump colonization losses in the residual stand. // Forest Service. Technical Bulletin № 1459. — 26 p.
- Garbelotto M. M., Lee H. K., Slaughter G., Popenuck T., Cobb F. W., Bruns T. D. Heterokaryosis Is Not Required for Virulence of Heterobasidion annosum. // Mycologia. — 1997. — V. 89 (1). — P. 92–102. DOI: 10.2307/3761177.
- Korhonen, K. and Stenlid, J. Biology of Heterobasidion annosum. Pages 43–71 in: Woodward S., Stenlid J., Karjalainen R., Hüttermann A. (eds) Heterobasidion annosum. Biology, Ecology, Impact and Control. — CAB International, 1998. — 589 p.
- Gibbs J. N. Resin and the resistance of conifers to Fomes annosus. // Annals of Botany. — 1968. — V. 32. — P. 649–665. DOI: 10.1093/oxfordjournals.aob.a084238.
- Morrison D. J., Redfern D. B. Long-term development of Heterobasidion annosum in basidiospore-infected Sitka spruce stumps. // Plant Pathology. — 1994. — V. 43 (5). — P. 897–906. DOI: 10.1111/j.1365-3059.1994.tb01634.x.
- Lygis V., Vasiliauskas R., Stenlid J. Planting Betula pendula on pine sites infested by Heterobasidion annosum: disease transfer, silvicultural evaluation and community of wood inhabiting fungi. // Canadian Journal of Forest Research. — 2004. — V. 34. — P. 120–130. DOI: 10.1139/x03-202.
- Lygis V., Vasiliauskas R., Stenlid J., Vasiliauskas A. Silvicultural and pathological evaluation of Scots pine afforestations mixed with deciduous trees to reduce the infections by Heterobasidion annosum. // Forest Ecology and Management. — 2004. — V. 201. — P. 275–285. DOI: 10.1016/j.foreco.2004.07.013.
- Stenlid J. Population structure of Heterobasidion annosum as determined by somatic incompatibility, sexual incompatibility, and isoenzyme patterns. // Canadian Journal of Botany. — 1985. — V. 63 (12). — P. 2268–2273. DOI: 10.1139/b85-322.
- Stenlid J.Controlling and predicting the spread of Heterobasidion annosum from infected stumps and trees of Picea abies. // Scandinavian Journal of Forest Research. — 1987. V. 2 (1–4). — P. 187–198. DOI: 10.1080/02827588709382457.
- Asiegbu F. O., Adomas A., Stenlid J. Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l. // Molecular plant pathology. — 2005. — V. 6 (4). — P. 395–409. DOI: 10.1111/J.1364-3703.2005.00295.X.
- Redfern D. B., Stenlid J. Spore dispersal and infection. Pages 105–125 in: Woodward S., Stenlid J., Karjalainen R., Hüttermann A. (eds) Heterobasidion annosum. Biology, Ecology, Impact and Control. — CAB International, 1998. — 589 p.
- Shaw C. G., Florance E. R. Scanning electron microscopy reveals differences in surface morphology between basidiospores and conidia of Heterobasidion annosum. // Forest Pathology. — 1979. — V. 9. — P. 249–254. DOI: 10.1111/J.1439-0329.1979.TB00685.X
- Moykkynen T. Liberation of Heterobasidion annosum conidia by airflow. // Forest Pathology. — 1997. — V. 27. — P. 283–289. DOI: 10.1111/J.1439-0329.1997.TB01082.X
- Hori C. et al. Analysis of the Phlebiopsis gigantea Genome, Transcriptome and Secretome Provides Insight into Its Pioneer Colonization Strategies of Wood. // PLOS Genetics. — 2014. — V. 10 (12). — P. 1–20. DOI: 10.1371/journal.pgen.1004759
- Звягинцев В. Б., Волченкова Г. А., Жданович С. А. Лесоводственные и лесозащитные мероприятия в пораженных корневой губкой сосновых насаждениях // Труды БГТУ. — 2013. — № 1. — С. 223–226.
- Roy G., Laflamme G., Bussieres G., Dessureault M. Field tests on biological control of Heterobasidion annosum by Phaeotheca dimorphospora in comparison with Phlebiopsis gigantea. // Forest Pathology. — 2003. — V. 33. — P. 127–140. DOI: 10.1046/j.1439-0329.2003.00319.x.
- Kallio T., Hallaksela A.-M. Biological control of Heterobasidion annosum (Fr.) Bref. (Fomes annosus) in Finland. // Forest Pathology. — 1979. — V. 9. — P. 298–308. DOI: 10.1111/j.1439-0329.1979.tb00692.x.
- Высоцкий А. А., Корчагин О. М. Корневая губка в насаждениях сосны обыкновенной (Pinus sylvestris L.). Проблемы и пути решения // Известия Санкт-Петербургской лесотехнической академии. — 2018. — Вып. 224. — С. 176–192. DOI: 10.21266/2079-4304.2018.224.176-192.
- Nikolajeva V. et al. Growth and antagonism of Trichoderma spp. and conifer pathogen Heterobasidion annosum s.l. in vitro at different temperatures. // Advances in Microbiology. — 2012. — V. 2 (3). — P. 295–302. DOI: 10.4236/aim.2012.23035.
- Anselmi N., Nicolotti G. Biological control of Heterobasidion annosum in the forest by non-pathogenic wood-destroying fungi. // Root and butt rots of forest trees: 9th International Conference on Root and Butt Rots, Carcans-Maubuisson, (France), 1–7 September, 1997. — 1998. — P. 421–428.
- Kenigsvalde K. et al. Impact of Trichoderma sp. on the development of Heterobasidion annosum in decayed understory Picea abies stumps. // Proceedings of the Latvian Academy of Sciences Section B Natural Exact and Applied Sciences. — 2017. — V. 71 (1–2). — P. 52–56. DOI: 10.1515/prolas-2017-0009.
- Korhonen K. Intersterility groups of Heterobasidion annosum. // Metsäntutkimuslaitoksen julkaisuja. — 1978. — V. 94 (6). — P. 1–25.
- Capretti P., Korhonen K., Mugnai L., Romagnoli C. An intersterility group of Heterobasidion annosum specialized to Abies alba. // Forest Pathology. — 1990. — V. 20 (4). — P. 231–240. DOI: 10.1111/j.1439-0329.1990.tb01134.x.
- Otrosina W. J., Garbelottob M. Heterobasidion occidentale sp. nov. and Heterobasidion irregulare nom. nov.: A disposition of North American Heterobasidion biological species. // Fungal biology. 2010. V. 114. — P. 16 – 25. DOI: 10.1016/j.mycres.2009.09.001.
- Yuan Y., Chen J.-J., Korhonen K., Martin F., Dai Y.-Ch. An Updated Global Species Diversity and Phylogeny in the Forest Pathogenic Genus Heterobasidion (Basidiomycota, Russulales). // Frontiers in Microbiology. — 2021. — V. 11. — P. 1–15. DOI: 10.3389/fmicb.2020.596393.
Ссылка на эту страницу для печатных изданий
Агеев Д. В., Бульонкова Т. М. Гетеробазидион многолетний (Heterobasidion annosum) – Грибы Сибири [Электронный ресурс] URL: https://mycology.su/heterobasidion-annosum.html (дата обращения: 29.05.2026).
Поделиться ссылкой
Дискуссии